Verbreitungsgebiete der verholzenden Päonienarten
Zur Herkunft der verholzenden Päonien (Strauchpfingstrosen)
Zentralchina (blau sind die wichtigsten Flüsse markiert, gelb die Städte, die für die Päonienkultur bekannt sind)
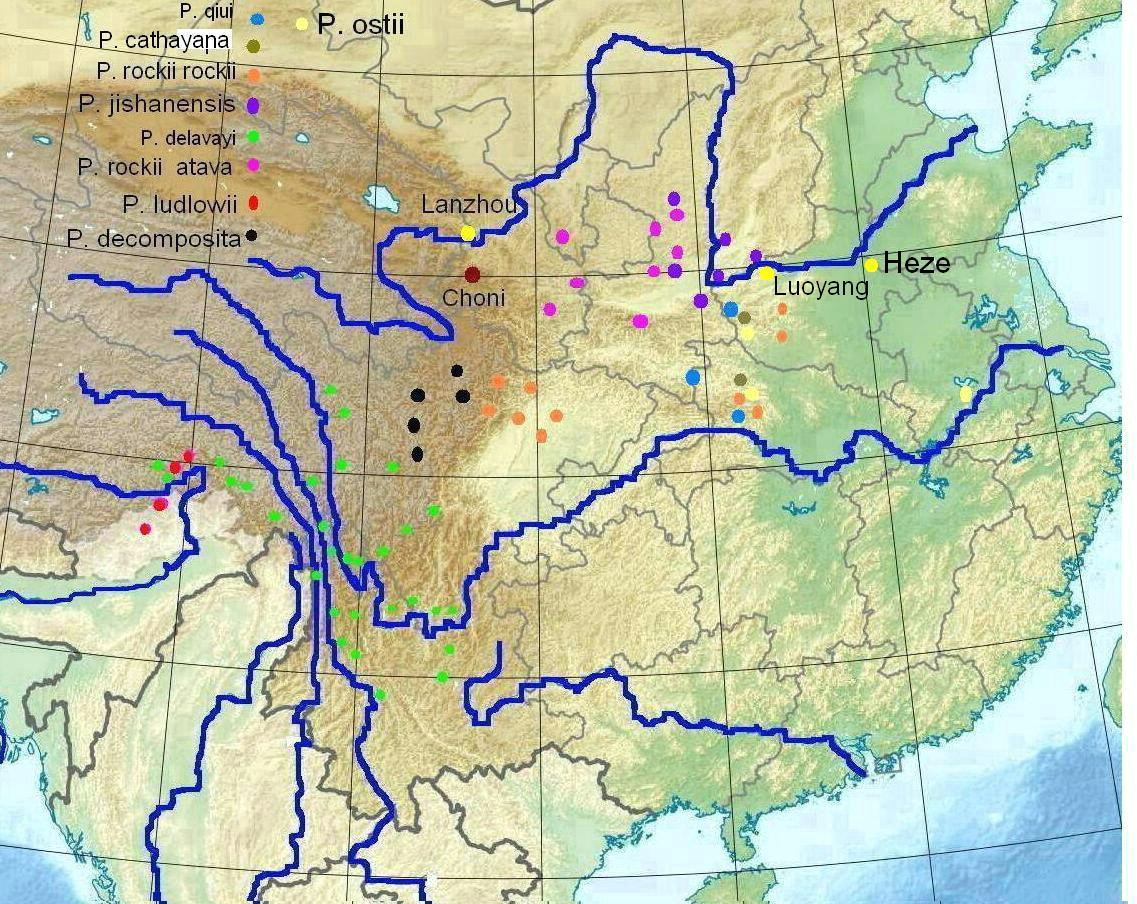
Nur in China kommen verholzende Päonien wild vor. Eine moderne und ausführliche Beschreibung der einzelnen Arten und ihrer Fundorte ist bei D.Y. HONG (2010) zu finden. Zu beachten ist, dass in der Päonienliteratur keine einheitliche botanische Bezeichnung der einzelnen Arten oder auch Unterarten vorliegt. In der beigefügten Darstellung sind wichtige Vorkommensorte in eine Karte von China eingetragen. Dabei werden für die Art P. rockii 2 Unterarten ausgewiesen: P. rockii ssp. rockii und P. rockii atava. Für die Unterart P. r. subsp. rockii wird von Päonienfreunden oft die veraltete (synonyme) Bezeichnung P. r. subsp. linyanshanii (T. HONG & G.L. OSTI ) verwendet. Dadurch können (sprachliche) Verwechslungen mit der Unterart atava vermieden werden.
Betrachtet man in der Darstellung die Fundorte für die beiden Unterarten von P. rockii, so fällt auf, dass sie sich nicht überlappen. Gemeinsam nehmen sie aber ein kompaktes Verbreitungsgebiet zwischen den Flüssen Jangtsekiang und Huang He ein. Am westlichen Randes dieses Verbreitungsgebietes befindet sich der Ort Choni mit dem Kloster, in dessen Garten J. Rock eine attraktive, ihm unbekannte Päonienart blühen sah – die später nach ihm benannte P. rockii. Dabei handelt es sich um die Unterart P. rockii subsp. atava. Zwischen den beiden P. rockii-Unterarten besteht keine Kreuzungsbariere. Dies spricht für ihre genetische Nähe. Die Fundpunkte für die kleinblütige , aber ähnliche Art P. decomposita schließen sich an das Verbreitungsgebiet von P. rockii an.
Für die heute als P. suffruticosa bezeichneten Kulturformen und Sorten kann keine Wildart ausgemacht werden. Es ist vorstellbar, dass die Wildarten P. cathayana, P. jishanensis, P. ostii und P. qiui an der Entstehung der ursprünglichen Formen der heutigen P. suffruticosa beteiligt waren. Unter den ersten in Europa eingeführten Sorten P. suffruticosa sind Formen zu finden, die über den für P. rockii typischen Basalfleck verfügen – ein Indiz dafür, dass auch diese Art zum Merkmalspool der heutigen P. suffrutiosa beigetragen hat. Zwischen den genannten Formen und Arten bestehen offensichtlich keine ausgeprägten Kreuzungsbarrieren. Dies deutet auf eine Artevolution in einem geschlossenen Verbreitungsgebiet hin.
Eine andere Situation liegt für die beiden Arten P. delavayi und P. ludlowii vor. Die Evolution dieser beiden Arten ist getrennt von der Evolution der oben genannten Arten erfolgt. Deswegen besteht eine stark ausgeprägte Kreuzungsbarriere zu den zentralchinesischen Formen. Dies wird durch die Geschichte der Entwicklung der Lutea-Hybriden (unter Verwendung von P. delavayi als Kreuzungspartner) anschaulich belegt. Über die Kreuzbarkeit von P. ludlovii z. B. mit P. rockii oder P. suffruticosa liegen keine Berichte vor.
Aus der beigefügten Abbildung (in Anlehnung an D.Y. HONG 2010)kann entnommen werden, dass die Evolution von P. delavayi in räumlich durch hohe Gebirgsketten getrennten Fluss-Systemen erfolgt ist. Dies kann ein Grund für die erhebliche Variabilität dieser Art sein, die taxonomisch erst durch HONG wieder zu einer Art vereint wurden. Die Evolution der beiden sehr ähnlichen Arten P. delavayi und P. ludlowii erfolgte vermutlich in keinem großen gemeinsamen Verbreitungsgebiet. Ungeachtet ihrer morphologischen Ähnlichkeit sind die beiden Arten reproduktiv voneinander getrennt. Diese Trennung ist allerdings nicht vollständig. Bei künstlicher Befruchtung können Bastarde zwischen P. ludlowii und P. delavayi erzielt werden. Nur in einem begrenzten Areal im Fluss-System des Bramaputra haben die beiden Arten ein sich überlappendes Verbreitungsgebiet.